本図鑑で対象とするサンゴ類
| 目次: ■有藻性サンゴ類 zooxanthellate corals ■刺胞動物門 Cnidaria ■花虫綱 Anthozoa ■六放サンゴ亜綱 Hexacorallia ■イシサンゴ目 Scleractinia ■イシサンゴ目内の亜目 ■八放サンゴ亜綱 Octocorallia ■ヒドロ虫綱 Hydrozoa ■分類体系の基準 ■引用文献ほか |
有藻性サンゴ類 zooxanthellate corals
 本図鑑で扱うサンゴ類は、有藻性サンゴ類 (zooxanthellate corals) である。本類は刺胞動物門 (表1:右) の中で、体内に褐虫藻 (図1) を共生させる種の総称で、花虫綱六放サンゴ亜綱イシサンゴ目のおよそ半数の種、花虫綱八放サンゴ亜綱硬八放サンゴ目アオサンゴ科と軟八放サンゴ目クダサンゴ科の全種、ならびにヒドロ虫綱花クラゲ目アナサンゴモドキ科の全種を含む。
本図鑑で扱うサンゴ類は、有藻性サンゴ類 (zooxanthellate corals) である。本類は刺胞動物門 (表1:右) の中で、体内に褐虫藻 (図1) を共生させる種の総称で、花虫綱六放サンゴ亜綱イシサンゴ目のおよそ半数の種、花虫綱八放サンゴ亜綱硬八放サンゴ目アオサンゴ科と軟八放サンゴ目クダサンゴ科の全種、ならびにヒドロ虫綱花クラゲ目アナサンゴモドキ科の全種を含む。
なお、有藻性サンゴ類に含まれる種は造礁サンゴ類 (hermatypic corals, reef-building corals) のそれとほぼ重なる。造礁サンゴ類は、褐虫藻を持ち、サンゴ礁形成に寄与する種の総称と定義される。しかしながら、褐虫藻を持っていてもサンゴ礁形成に関与しない種が存在し、また、褐虫藻を持っていない種が深海域にサンゴ礁を形成することがあるなど
(Roberts et al. 2009)、その定義には曖昧さがつきまとう。そのため、サンゴ類研究においては、「造礁サンゴ類」の代わりに定義がより厳密な「有藻性サンゴ類」が用語として使われるようになってきている。
以下に、本図鑑の主対象である有藻性サンゴ類が属する分類群について、簡単に解説する。表1を併せて参照されたい。なお、本項を執筆するにあたり参考にした文献を末尾に記した。また、必要に応じて本文中に引用文献を記した。
 1
1
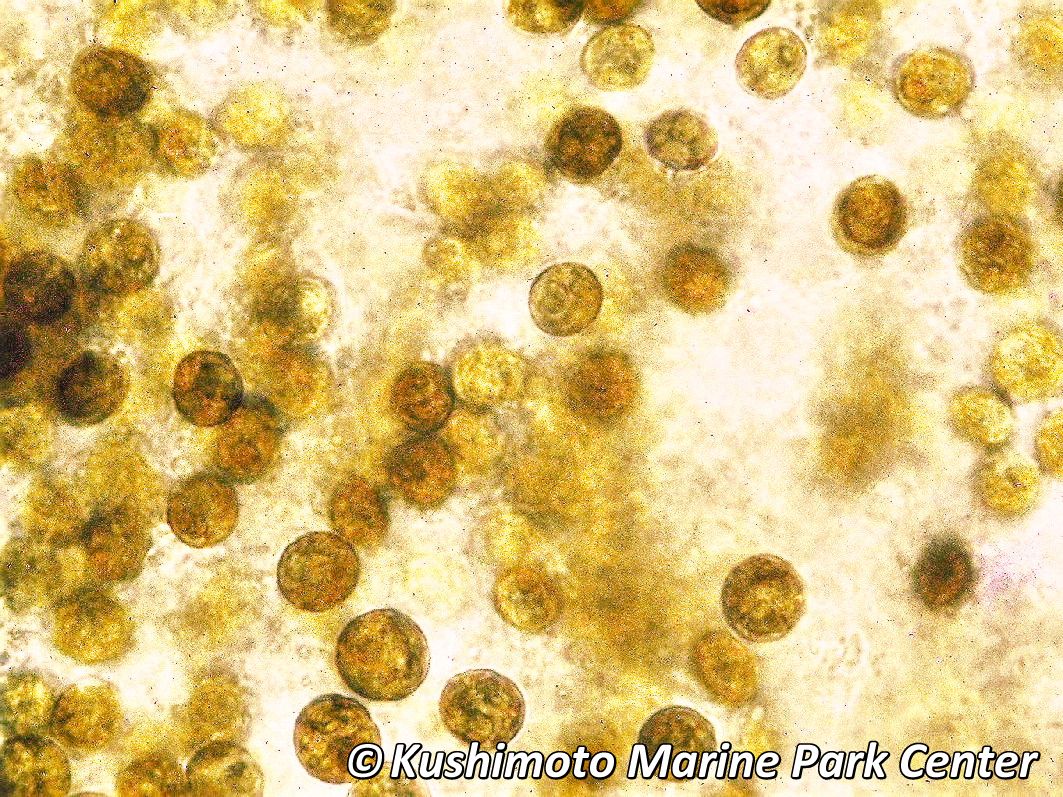
図1. 褐虫藻 (zooxanthellae). 渦鞭毛藻類 (うずべんもうそう) に属する単細胞藻類の総称で、褐色で共生状態ではほぼ球形を成し、直径はおよそ 10 µm (1 mm の 100分の1)。光合成栄養物を宿主に与え、また、宿主の色素としても働く。(写真:串本海中公園センター)
刺胞動物門 Cnidaria
刺胞動物門の祖先は先カンブリア紀に出現し、世界で現生種は10,203種、化石種は6,180種がそれぞれ知られる (Zhang 2013)。6綱に分かれ、放射相称もしくは左右相称の二胚葉動物
(外胚葉と内胚葉の2種の体制を持つ動物) で、体腔を欠き、体内の唯一の腔所である胃腔は消化と排泄を兼ね (口と肛門が一緒)、「刺胞」と呼ばれる細胞内小器官を持つ
(図2)。ヒドロ虫綱のごく一部を除いて海産で、漂泳性 (クラゲ) と付着性 (ポリプ) ならびに単体性と群体性の2つの異なる生活様式があり、他の動物と共生関係を持つものも少なくない。
 2
2
図2. 刺胞 (nematocyst). 毒針の入った細長い袋状の細胞内小器官で、長さは大きいもので数 100 µm ある。刺激を受けると袋を反転させて毒針が自動発射され、捕食や防御用の武器として用いられる。(写真:串本海中公園センター)
花虫綱 Anthozoa
花虫綱は以下に記す他の刺胞動物に見られない特徴を持つ:一生をポリプ世代で過ごしクラゲ世代を欠く、口道から胃腔内部を放射状に仕切る膜 (隔膜)
が6枚以上ある、口は円形ではなくてスリット型である、口と胃腔の間に外胚葉性の口道を持つ。
花虫綱はハナギンチャク亜綱、六放サンゴ亜綱、八放サンゴ亜綱の3亜綱に分かれる。ハナギンチャク亜綱はイソギンチャクの様な形をしているが、固着せず、砂地に掘った穴内に粘液で棲管を作り生活する。内部形態は六放サンゴ亜綱に似るが、特異な幼生型を持つ。八放サンゴ亜綱はポリプの触手の数は常に8本で、触手の両側に羽状突起を備え、隔膜も常に8枚である。
六放サンゴ亜綱 Hexacorallia
ポリプは常に同形。イソギンチャク目、ツノサンゴ目、ホネナシサンゴ目、イシサンゴ目、スナギンチャク目の5つの目で構成される。六放サンゴ亜綱は八放サンゴ亜綱に比べて内包する分類群のまとまりが悪く、触手や隔膜の数は分類群によって異なり、6、10、12
またはそれらの倍数を基本としている。また、口道には管溝を有するが、その数も分類群によって異なる。
イソギンチャク目は全て単体性で骨格を欠き、多くは大型のポリプを持ち、たいてい足盤で他物に固着する。隔膜と触手の数は12以上である。
ツノサンゴ目は角質の固い骨軸から成る樹木状の群体を形成し、一部の大型種の骨軸は黒珊瑚と呼ばれ、装飾品として利用される。骨軸の周りを薄い肉部が包み、骨軸にはポリプが入る穴はない。骨軸上には小さな棘が無数にあり、角珊瑚の名前の由来になっている。触手の数は6本、隔膜の数は6~12枚である。
ホネナシサンゴ目は単体性もしくは群体性で骨格を欠く。形はイソギンチャク目に似るが、足盤を欠き、基質にしっかりと固着するため、はがすことは容易ではない。隔膜と触手の数は12以上で、体の構造はイソギンチャク目よりはイシサンゴ目に近い。
スナギンチャク目は骨格を欠き、多くの種が共肉で連なった群体性であるが、単体性の種もある。隔膜と触手の数は12以上である。名前は一部の種が体壁中に砂粒等の異物を取り込むことがあることに由来する。
イシサンゴ目 Scleractinia
個体 (individual) の基本体制は、ポリプ (polyp、個虫とも呼ばれる) に関連する軟体部 (soft body) と、石灰質(主に炭酸カルシウム)の骨格
(サンゴ個体:corallite) から構成される。軟体部はさらにポリプと共肉 (coenosarc) より構成される (図3)。個体とサンゴ個体は紛らわしいが、後者は前者の骨格部分を指す。また、個体の集合体は群体
(colony:図4)、その骨格部分はサンゴ体 (corallum:図5) と呼ばれる。
 3
3  4
4  5
5
図3. 個体の基本体制. 西平(1991)を参考にして描く.
図4. オオスリバチサンゴ Duncanopsammia peltata の群体.
図5. オオスリバチサンゴ Duncanopsammia peltata のサンゴ体.
図はいずれも野村恵一.
ポリプは骨格に空いた莢 (きょう:calice) と呼ばれる穴内に収まる。サンゴ個体の莢内には、莢の中心に向かって放射状に配列する薄板状もしくは鋸歯状の構造物が見られ隔壁
(septum) と呼ばれる。隔壁は一般に6枚の1次隔壁の間に6枚の2次隔壁が生じ、これらの間に12枚の3次隔壁、同様に24枚の4次隔壁、48枚の5次隔壁と続くが、分類群によっては2次隔壁以降を持たないものや、次数の判別が難しいもの、ハナガササンゴ型配列やキサンゴ型配列のように特殊な形態を示すものがある。隔壁の上縁は鋸歯状をなす場合が多く、最内縁の他よりも大きくて葉状のものはパリ状葉
(paliform lobe) と呼ばれる。隔壁の内側、すなわち莢の中央にはたいてい針状、杭状、板状もしくはスポンジ状の構造物が見られ軸柱 (columella)
と呼ばれる。莢の外周部分は莢壁 (theca) と呼ばれるが、莢壁と共骨の区別が明瞭でない場合は総称して個体壁 (corallite wall)
と呼ばれる場合がある。隔壁から莢壁の外側に伸びた畝状の部分は肋 (costa) と呼ばれる (図6)。ポリプは、自在に伸長したり莢の内部に退縮することが出来る。伸長時は触手
(tentacle) を広げ、その数は12本以上である。触手の根元の内側には口盤 (oral disc) とよばれる裸地部があり、その中央に口が位置する。口と胃腔
(gastric cavity) の間には口道 (stomodaeum) があり、口道や胃腔は12枚以上の隔膜 (mesentery) の襞に被われる。胃腔の壁面には隔膜の一部に包まれた生殖腺
(gonad) があり、隔膜の下部には隔膜糸 (mesenterial filament) と呼ばれる消化器官がある (図3)。食べたものは胃腔内で消化され糞は口から排出される。
 6
6
図6. キクメイシの一種 Dipsastraea sp. のサンゴ個体の構造.
イシサンゴ目の分類は、主に上述したサンゴ個体やサンゴ体の構造に基づいて行われるが、明瞭な分類形質が乏しい上に、種内形態変異が著しく、形態のみでの分類には限界があった。そのため、近年は分子系統解析に基づいたイシサンゴ目の分類体系の大幅な改変が進んでいる
(例えば、Fukami et al. 2008, Gittenberger et al. 2011, Benzoni et al. 2012, Budd et al. 2012, Huang et al. 2014, Huang et al. 2016)。
※参考:日本産イシサンゴ目分類体系の新旧比較
 現在、イシサンゴ目は36科 (表2:右) にに分けられ (WoRMS 2021)、この内の日本産は30科で、Astrangiidae, Cladocoridae, Faviidae,
Gardineriidae, Meandrinidae, Montastraeidae の6科は日本から知られていない。また、褐虫藻との共生の有無は科ごとにほぼまとまっており、18科は全ての種が有藻性、13科は全ての種が無藻性で、5科
(ヒラフキサンゴ科、チョウジガイ科、キサンゴ科、ハナヤサイサンゴ科、シオガマサンゴ科) には有藻性の種と無藻性の種の両方が含まれる (表2)。イシサンゴ目に属する現生種の種数は1,500以上と推定され
(Madin et al. 2016)、有藻性イシサンゴ類と無藻性イシサンゴ類のそれぞれの構成種数はほぼ半々とされている (Cairns 1994, Roberts et al. 2009)。
現在、イシサンゴ目は36科 (表2:右) にに分けられ (WoRMS 2021)、この内の日本産は30科で、Astrangiidae, Cladocoridae, Faviidae,
Gardineriidae, Meandrinidae, Montastraeidae の6科は日本から知られていない。また、褐虫藻との共生の有無は科ごとにほぼまとまっており、18科は全ての種が有藻性、13科は全ての種が無藻性で、5科
(ヒラフキサンゴ科、チョウジガイ科、キサンゴ科、ハナヤサイサンゴ科、シオガマサンゴ科) には有藻性の種と無藻性の種の両方が含まれる (表2)。イシサンゴ目に属する現生種の種数は1,500以上と推定され
(Madin et al. 2016)、有藻性イシサンゴ類と無藻性イシサンゴ類のそれぞれの構成種数はほぼ半々とされている (Cairns 1994, Roberts et al. 2009)。
イシサンゴ目には群体性と単体性、ならびに固着性と非固着性のそれぞれ2つの生活様式があり、また、繁殖方法も有性生殖と無性生殖の2つの様式がある。さらに、褐虫藻を持つ有藻性イシサンゴ類と褐虫藻を持たない無藻性イシサンゴ類の2つに生態学的に分類され、前者は褐虫藻の光合成に日光が必要なため基本的に浅所に生息し、生育に日光を必要としない後者は深所に多いが浅所に生息する種も見られる。
イシサンゴ目内の亜目
イシサンゴ目の分類体系は、形態学的解析によって Vaughan & Wells (1943) により5つの亜目 (ムカシサンゴ亜目 Astrocoeniina、クサビライシ亜目
Fungiina、キクメイシ亜目 Faviina、チョウジガイ亜目 Caryophylliina、キサンゴ亜目 Dendrophylliina)
に分類され、後に Alloiteau (1952) により8亜目に、Chevalier & Beauvais (1987) により11亜目に改変された。しかしながら、一般的には2000年代初めまで基本的に
Vaughan & Wells (1943) の分類体系が踏襲されてきた。ところが、1990年代以降、分子系統解析により、従来とは全く異なる分類体系が示されるようなり、Romano
& Palumbi (1996) はイシサンゴ目を Complex と Robust の2つのクレードに大別し、Stolarski et al. (2011) はこれに加えて深所性無藻性イシサンゴ類の一部に Basal という第三のクレードの存在を示した。
一方で、Okubo (2016) はイシサンゴ目を個体の発生様式により2つに大別し、それらは Complex と Robust クレードに相当することを突き止め、これら胚の発生様式と分子系統の両面に基づいてイシサンゴ目に2つの亜目
(シズカテマリ亜目 Refertina とナミフウセン亜目 Vacatina) を提唱した。シズカテマリ亜目は、胚の発生過程で胞胚腔を欠くかほとんど見られず、分子系統では
Complex クレードに相当する。ナミフウセン亜目は、胚の発生過程で明らかな胞胚腔が見られ、分子系統では Robust クレードに相当する
(Okubo 2016, 大久保 2016)。
ただし、無藻性イシサンゴ類の多くの種においては発生様式の検討が不十分であるため (特に分子系統の Basal クレード)、WoRMS (2021)
では Okubo (2016) により提唱された2つの亜目は要再検討 (taxon inquirendum) としている。このように、イシサンゴ目全体にわたる亜目の解析は十分には成されていないが、本図鑑で扱う有藻性イシサンゴ類においては解析が十分に進んでおり、かつ将来的にも普遍性のある分類単位であると判断されたため、本図鑑においては
Okubo (2016) の2つの亜目を採用している。
※Okubo (2016) が提唱した2亜目は、2023年12月9日、WoRMS (2023) において有効名とされた。
八放サンゴ亜綱 Octocorallia
大多数の種が摂食に関与する通常ポリプのみを持つが、触手を持たない管状ポリプ等の多型ポリプを持つ種もある。管状ポリプに生殖機能を持つ種もある。通常ポリプの触手と隔膜の数は常に8、多くの種において触手に羽肢を持つことが特徴であり、六放サンゴ亜綱と比して非常にまとまった分類群である。一部の種を除き群体性、共肉には微小骨片を有する種が大勢である。また、群体を支える石灰質や角質の骨軸を形成する種もあり、ごく少数イシサンゴ目に類似の骨格を形成する種もある。
昨今、系統分類学においても遺伝子情報に基づいた研究が進んでおり、McFadden et al. (2022) は従来のウミトサカ目・アオサンゴ目・ウミエラ目という形態を基本とした分類体系を廃し、分子系統解析により分かれた2つのクレードをそれぞれ
Malacalcyonacea 軟八放サンゴ目と Scleralcyonacea 硬八放サンゴ目の2目とした。
軟八放サンゴ目
独特の管状骨格を有するクダサンゴはこの目に属する。直径数ミリのポリプには羽肢を持った8本の触手が観察できる。その他、浅海サンゴ礁域で多く見られるウミキノコ属、ウネタケ属、カタトサカ属の各種は有藻性で成長も速く、場所によっては大群落を形成する。共肉は微細な石灰質の骨片が充満しており、これで群体全体を支えている。カタトサカ属は肉眼でも確認できる長さ数ミリの巨大な骨片を持ち、サンゴ礁形成にも関与している
(Jeng et al. 2011)。礁斜面に生息するイソバナ属やヒラヤギ属などは、赤やオレンジの目立つ色彩と直径 1 m 以上の扇状群体に成長する。
硬八放サンゴ目
この目にはサンゴ礁域に生息するアオサンゴが含まれる。アオサンゴはイシサンゴ目に類似の堅固な骨格を持ち、その群体形もよく似るが、ポリプには8本の触手を持つ。砂泥地に生息するウミエラ・ウミサボテン類には有藻性の種もあり、礁池内の砂地で群生することもある。その堅固な骨軸が宝飾品として利用される、いわゆる「宝石サンゴ」もこの目に属する。
ヒドロ虫綱 Hydrozoa
ヒドロ虫綱は、口道や隔膜を欠くなどの点で他の刺胞動物と異なる。多くの種でポリプ世代とクラゲ世代の世代交代を行う。群体性種と単体性種の両方があり、群体性種では個虫の機能分化し
(カツオノエボシなど機能分化が著しい種もある)、口と消化器を有する栄養個虫と口と消化器を欠く指状個虫の2型がある。繁殖期には生殖体が栄養個虫上またはヒドロ根上に形成され、クラゲを産する。
ヒドロ虫綱は、Anthoathecata (シノニムとして Anthomedusae 花クラゲ目)、Leptothecata (シノニムとして
Leptomedusae 軟クラゲ目)、Siphonophorae クダクラゲ目、Actinulida アクチヌラ目、Limnomedusae
淡水クラゲ目、Narcomedusae 剛クラゲ目、Trachymedusae 硬クラゲ目の6目で構成される (Japan Agency for
Marine-Earth Science and Technology 2023, Schuchert 2023)。Schuchert (2023)
によると、ヒドロ虫綱は刺胞動物門のなかでも最も多様性が高いグループで、現生種は約3,800種にのぼるが (Zhang (2013) が示した刺胞動物門6綱合計の10,203種に対して約37%に相当する)、多数のシノニム種が含まれていると想定されるほか、分類体系、特に上位の分類階級については広くコンセンサスが得られていないという。
ヒドロ虫綱で骨格を形成するのは Anthoathecata に属する Milleporidae アナサンゴモドキ科 と Stylasteridae
サンゴモドキ科で、うち有藻性であるのはアナサンゴモドキ科のみである。この科は Millepora アナサンゴモドキ属のみで構成され、World Hydrozoa Database には 17種2亜種が有効名として挙げられているほか、11変種
(variety)、5型 (forma) も併記されている (Schuchert 2023)。Takama et al. (2017) は、サンゴモドキの5種についてITSクレードを調べたところ形態に基づく分類とは一致しないことから、分類体系の見直しの必要性を指摘している。
分類体系の基準
有藻性サンゴ類の分類体系は、2000年代以降、DNA ならびにミクロな骨格形態の情報を取り入れることによって大幅に見直されてきており(Stefani
et al. 2007, Fukami et al. 2008, Benzoni et al. 2010, 2014, Gittenberger et al. 2011, Budd et al. 2012, Kitano et al. 2014, Huang et al. 2016, Luzon et al. 2017, Oku et al. 2017 など)、その見直しは現在も続けられ、系統関係の再構築、シノニム関係の整理、新分類群や新種等の記載が頻繁に行われている。
この図鑑における分類体系は、可能な限り原記載ならびに担名タイプを確認した上で*1、個別の論文等*2を確認したほか、海洋生物の分類データーベースである WoRMS*3も参照して記述した。ただし、論文や WoRMS の情報に誤りや疑問がある場合、次のとおり扱った。
①反論根拠があれば、その根拠を明示して WoRMS と異なる扱いを行う。
②反論根拠を示せなければ、WoRMS and/or 出典文献の不備、疑問点や今後の課題などを列記する。
③誤記があれば根拠を示して訂正する。
また、執筆者らの研究に基づき、別種、隠蔽種、未記載種あるいはその可能性があると判断した場合は、注記を付して採録した。分類学的位置が未確定であるが、未記載種である可能性がもたれるものについては
sp. に続き便宜的な識別番号や識別名を与えた。
脚注:
*1. 原記載や担名タイプが確認できない種や分類階級も存在する。また、日本語・英語以外の言語で書かれた資料については、語学能力的問題から、その内容を正確に理解するのが難しいケースもある。
*2. 国際動物命名規約(日本分類学連合会)3章および4章に準じる「公表された出版物」に該当すれば、根拠論文としての条件を満たすと判断した。
Rowlett (2020) では、分類体系の変更やシノニムの扱いなどについて明確な根拠が示されていない記述が多く見られるが、WoRMS などでは根拠論文の一つとして扱われている。Rowlett
(2020) に限らず既刊の図鑑等は、学会誌と同様の査読を受けていることはほとんどないと思われるが、分類の根拠資料として研究者に広く受け入れられているのは事実である。それも踏まえると、国際動物命名規約の3章および4章に準じる「最後発の公表された出版物」であれば、査読を経た論文でないことを理由に無視することは適当でない
(ただし内容の妥当性は別問題である)。
*3. WoRMS: World Register of Marine Speciesの略で、国際的な海洋生物データベース。 イシサンゴ目ては World List of Scleractinia 、八放サンゴ亜綱は World List of Octocorallia、ヒドロ虫綱は World Hydrozoa Database の情報が参照掲示されている。
WoRMS は海洋生物データベースのデファクトスタンダードになっていることから、この図鑑における分類体系は、WoRMS の分類体系との整合性に注意を払うこととした。これは、異なる分類ポリシーによる複数の分類体系が並立する弊害は大きいためである。ただし、反論根拠がある場合は、その根拠を明示して
WoRMS と異なる扱いを行った。
引用文献ほか
引用文献:
Alloiteau J (1952) Madréporaires post-paléozoiques. In: Piveteau J (ed) Traité de paléontologie. Masson, Paris, pp 539-684.
Benzoni F, Arrigoni R, Stefani F, Stolarski J (2012) Systematics of the coral genus Craterastrea (Cnidaria, Anthozoa, Scleractinia) and description of a new family through combined morphological and molecular analyses. System Biodivers 10: 417-433. [ResearchGate]
Benzoni F, Arrigoni R, Waheed Z, Stefani F, Hoeksema BW (2014) Phylogenetic relationships and revision of the genus Blastomussa (Cnidaria: Anthozoa: Scleractinia) with description of a new species. Raffles Bulletin of Zoology, 62: 358–378. [ResearchGate]
Benzoni F, Stefani F, Pichon M, Galli P (2010) The name game: Morpho-molecular species boundaries in the genus Psammocora (Cnidaria, Scleractinia). Zool J Linn Soc 160: 421–456.
Budd AF, Fukami H, Smith ND, Knowlton N (2012) Taxonomic classification of the reef coral family Mussidae (Cnidaria: Anthozoa: Scleractinia). Zool J Linn Soc 166: 465-529. [Wiley Online Library]
Cairns SD (1994) Scleractinia of the temperate north Pacific. Smithsonian Contributions to Zoology, 557. Smithson Ianinstitution Press, Washington, DC. [Smithsonian Research Online]
Chevalier JP, Beauvais L (1987) Ordre des scleractiniaires. In: Grasse PP (ed) Traite de zoologie, Cnidaires, Anthozoaires. Vol. 3. Masson, Paris, pp 403-764.
Fukami H, Chen CA, Budd AF, Collins A, Wallace C, Chuang Y, Chen C, Dai C, Iwao K, Sheppard C, Knowlton N (2008) Mitochondrial and nuclear genes suggest that stony corals are monophyletic but most families of stony corals are not (order Scleractinia, class Anthozoa, phylum Cnidaria). PLoS ONE 3: e3222. [PLoS ONE]
Gittenberger A, Reijnen BT, Hoeksema BW (2011) A molecularly based phylogeny reconstruction of mushroom corals (Scleractinia: Fungiidae) with taxonomic consequences and evolutionary implications for life history traits. Contrib Zool 80: 107-132. [ResearchGate]
Huang D, Arrigoni R, Benzoni F, Fukami H, Knowlton N, Smith ND, Stolarski J, Chou LM, Budd AF (2016) Taxonomic classification of the reef coral family Lobophylliidae (Cnidaria: Anthozoa: Scleractinia). Zool J Linn Soc 178: 436-481. [Wiley Online Library]
Huang D, Benzoni F, Fukami H, Knowlton N, Smith ND, Budd AF (2014) Taxonomic classification of the reef coral families Merulinidae, Montastraeidae, and Diploastraeidae (Cnidaria: Anthozoa: Scleractinia). Zool J Linn Soc 171: 277-355. [Wiley Online Library]
Japan Agency for Marine-Earth Science and Technology (2023) Hydrozoa Owen, 1843. Biological Information System for Marine Life at https://www.godac.jamstec.go.jp/bismal/j/view/0000068. Accessed on 2023-10-02.
Jeng MS, Huang HD, Dai CF, Hsiao YC, Benayahu Y(2011)Sclerite calcification and reef-building in the fleshy octocoral genus Sinularia (Octocorallia: Alcyonacea). Coral Reefs 30: 925–933 DOI 10.1007/s00338-011-0765-z.
Kitano YF, Benzoni F, Arrigoni R, Shirayama Y, Wallace CC, Fukami H (2014) A phylogeny of the family Poritidae (Cnidaria, Scleractinia) based on molecular and morphological analyses. PLoS ONE 9: e98406. [PLoS ONE]
Luzon KS, Lin MF, Lagman MCAA, Licuanan WRY, Chen CA (2018) Correction: Resurrecting a subgenus to genus: molecular phylogeny of Euphyllia and Fimbriaphyllia (order Scleractinia; family Euphylliidae; clade V). PeerJ 6: e4074/correction-1. [PeerJ]
Madin JS, Anderson KD, Andreasen MH, Bridge TCL, Cairns SD, Connolly SR, Darling, ES, Diaz M, Falste DS, Franklin EC, Gates RD, Harmer AMT, Hoogenboom MO, Huang D, Keith SA, Kosnik MA, Kuo C-Y, Lough JM, Lovelock CE, Luiz O, Martinelli J, Mizerek T, Pandolfi JM, Pochon X, Pratchett MS, Putnam HM, Roberts TE, Stat M, Wallace CC, Widman E, Baird AH (2016) The Coral Trait Database, a curated database of trait information for coral species from the global oceans. Sci Data, 3: 160017. [Scientific Data]
McFadden CS, van Ofwegen LP, Quattrini AM (2022) Revisionary systematics of Octocorallia (Cnidaria: Anthozoa) guided by phylogenomics. Bull Soc Sys Biol 1: 8735. 79pp. doi.org/10.18061/bssb.v1i3.8735.
西平守孝 (1991) フィールド図鑑 造礁サンゴ 増補版. 東海大学出版会, 東京.
西平守孝・Veron JEN (1995) 日本の造礁サンゴ類. 海游社, 東京.
Oku Y, Naruse T, Fukami H (2017) Morpho-molecular evidence for polymorphism in the mushroom coral Cycloseris hexagonalis (Scleractinia: Fungiidae), with a new phylogenetic position and the establishment of a new genus for the species. Zool Sci 34: 242-251. [ResearchGate]
Okubo N (2016) Restructuring the Traditional Suborders in the Order Scleractinia Based on Embryogenetic Morphological Characteristics. Zool Sci 33: 116-123. [BioOne]
大久保奈弥 (2016) 発生 多様なサンゴの発生観察からようやく見えてきた共通性. 生物科学 68: 11-23.
Roberts JM, Wheeler A, Freiwald A, Cairns S (2009) Cold-water corals. The biology and geology of deep-sea coral habitats. Cambridge University Press, Cambridge.
Romano SL, Palumbi SR (1996) Evolution of scleractinian corals inferred from molecular systematics. Science 271: 640-642. [ResearchGate]
Rowlett J (2020) Indo-Pacific corals. Rowlett (self-published). [WoRMS]
Schuchert P (2023) World Hydrozoa Database. Accessed at https://www.marinespecies.org/hydrozoa on 2023-10-01. doi:10.14284/357.
Stefani F, Benzoni F, Pichon M, Cancelliere C, Galli P (2007) A multidisciplinary approach to the definition of species boundaries in branching species of the coral genus Psammocora (Cnidaria, Scleractinia) Zool Scr 37: 71-91. [ResearchGate]
Stolarski J, Kitahara MV, Miller DJ, Cairns SD, Mazur M, Meibom A (2011) The ancient evolutionary origins of Scleractinia revealed by azooxanthellate corals. BMC Evol Biol 11: 316. [BioMedCentral]
Takama O, Fernandez-Silva I, López C, Reimer JD (2018) Molecular phylogeny demonstrates the need for taxonomic reconsideration of species diversity of the hydrocoral genus Millepora (Cnidaria: Hydrozoa) in the Pacific. Zoological Science 3: 123-133. [BioOne]
Vaughan TW, Wells JW (1943) Revision of the suborders, families and genera of the scleractinia. Special papers of the Geological Society of America 44. Waverly Press, Baltimore, MD. [GeoScienceWorld]
WoRMS (2021) Cnidaria. Accessed at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=1267 on 2021-07-13.
WoRMS (2023). Scleractinia. Accessed at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=1363 on 2023-12-9
WoRMS (2023) Octocorallia. Accessed at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=1341 on 2023-10-02.
Zhang ZQ (2013) Animal biodiversity: An update of classification and diversity
in 2013. Zootaxa 3703: 5-11. [Zootaxa]
参考文献等:
美ら島財団・黒潮生物研究所 (2016) 美ら島研究センター収蔵資料目録 沖縄のソフトコーラル. 美ら島財団・黒潮生物研究所, 本部町・大月町. [沖縄美ら島財団]
江口元起 (1968) 相模湾産ヒドロ珊瑚類および石珊瑚類. 丸善, 東京.
今原幸光 (1992) クダサンゴ. In: 西村三郎 (編著) 原色検索日本海岸動物図鑑Ⅰ. 保育社, 大阪, pp 73-74, 図版13-4, 5.
久保田信 (2000) 刺胞動物門. In: 白山義久 (編著) 無脊椎動物の多様性と系統 (節足動物を除く). 裳華房, 東京, pp 108-112.
峯水亮・久保田信・平野弥生・ドゥーグル・リンズィー (2015) 日本クラゲ大図鑑. 平凡社, 東京.
西村三郎・久保田信・今原幸光・岩瀬文人・内田紘臣・酒井一彦 (1992) 刺胞動物門. In: 西村三郎 (編著) 原色検索日本海岸動物図鑑Ⅰ. 保育社, 大阪, pp 20-167.
小川数也 (2006) 相模灘海域のイシサンゴ類相. 国立科学博物館専報 40: 103-112.
Stolarski J, Roniewicz E (2001) Towards a new synthesis of evolutionary relationships and classification of Scleractinia. J Paleontol 75: 1090-1108. [ResearchGate]
Veron JEN (2000) Corals of the world. Australian Institute of Marine Science, Townsville.
座安佑奈・横地洋之・梶原健次・木村匡・島田剛・下池和幸・鈴木豪・立川浩之・長田智史・野村恵一 (2017) イシサンゴ分類の現状と日本造礁サンゴ分類研究会の取組. タクサ 42: 10–15. [J-Stage]
──────────
執筆者:野村恵一・横地洋之・立川浩之・野中正法・梶原健次
Citation:
更新履歴:
2023-11-12 公開
2023-12-09 イシサンゴ目の項に日本産イシサンゴ目分類体系の新旧比較のリンクを追加。
2023-12-12 WoRMS にて要再検討 (taxon inquirendum) となっていたイシサンゴ目の2亜目が有効とされたことを受け、「イシサンゴ目内の亜目」の項に注記を追加。
2025-08-04 イシサンゴ目の項中、日本産科に Stenocyathidae を追加するなど科数に関する記述を修正。