Scleractinia イシサンゴ目
Fungiidae Dana, 1846
クサビライシ科
本科に含まれる日本産属
Ctenactis Verrill, 1864 トゲクサビライシ属
Cycloseris Milne Edwards & Haime, 1849 マンジュウイシ属
Danafungia Wells, 1966 ナミトゲクサビライシ属
Fungia Lamarck, 1801 シタザラクサビライシ属
Halomitra Dana, 1846 カブトサンゴ属
Heliofungia Wells, 1966 パラオクサビライシ属
Herpolitha Eschscholtz, 1825 キュウリイシ属
Lithophyllon Rehberg, 1892 カワラサンゴ属
Lobactis Verrill, 1864 クサビライシ属
Pleuractis Verrill, 1864 ゾウリイシ属
Podabacia Milne Edwards & Haime, 1849 ヤエヤマカワラサンゴ属
Polyphyllia Blainville, 1830 ナマコイシ属
Sandalolitha Quelch, 1884 ヘルメットイシ属
Sinuorota Oku, Naruse & Fukami, 2017 ハグルマサンゴ属
Zoopilus Dana, 1846 アミガササンゴ属
本科の解説
解説: 解説:有藻性で単口性または多口性。多くの種で成体は自由生活性であるが、一部の種は成体となっても固着性である。本科の生活環については生活環の項で詳述するが、概要は以下のとおりである。有性生殖により形成されたプラヌラ幼生が着底すると、マツタケやシイタケのようなキノコに類似した形態の単口性・固着性の芽体が形成される。自由生活性の種では、芽体が成長するとキノコの傘にあたる部分
(口とその周辺の骨格) が自切 (横分裂) によって柄部から分離し、自由生活体になる。自由生活体は海底に横たわった状態で成長し、成熟すると有性生殖を行う。固着性の種では横分裂が起こらず固着した状態で成長し成熟する。本科の有性生殖様式は十分に研究されていないが、報告されている種は雌雄異体・放卵放精またはプラヌラ放出型で、雄性先熟の性転換をする種も知られている
(Loya et al. 2009)。
芽体に最初に形成される口 (初口) は一つ (単口) であるが、口内分裂または口周縁出芽(いずれも触手環内出芽に含まれる)により初口以外の口が二次的に形成されると多口性となる。成長すると例外なく多口性となる種と、成長しても基本的に単口性のままの種があるが、後者においても二次的な口が形成され複数の口を持つようになったサンゴ体がしばしば観察される。
なお、Hoeksema (1989)は、口が一つのサンゴ体について単体性 (solitary) ではなく単口性 (monostomatous
または monocentric)、口が複数のサンゴ体に群体性 (colonial) ではなく多口性 (polystonatous または polycentric)
の用語を用いている。これは colony には群体以外に個体の集合の意味もあり、自由生活性のサンゴ体の集合も colony との表現が可能であることから、colony–colonial
の用語を避けたためと思われる。本図鑑のクサビライシ科に関する記述では、Hoeksema (1989) に倣い単口性・多口性の用語を用いる。
単口で自由生活性の種は通常円形~楕円形の円盤状で、上面が平面状または盛り上がり、下面が平面状またはやや凹む。サンゴ体は、上面中央付近の口を中心として放射状に配列する隔壁と、それに対応する位置にある肋が一枚の板状のユニット
(隔壁-肋ユニット) となり、隣接するユニット間は複合シナプティキュラおよび底面付近に位置するサンゴ体壁で結合され、全体として円盤状のサンゴ体が形成される
(図1)。このため、多くのイシサンゴ類に見られる莢の周縁を囲む莢壁を欠く。なお、クサビライシ科では肋はサンゴ体の下面に形成される放射状の隆起を指し、上面全体に放射状に配列する板状の骨格を隔壁と呼ぶ。このため、多口性のサンゴ体で隣接する口の間を結ぶ板状の骨格は、肋ではなく口間隔壁と呼ばれる。
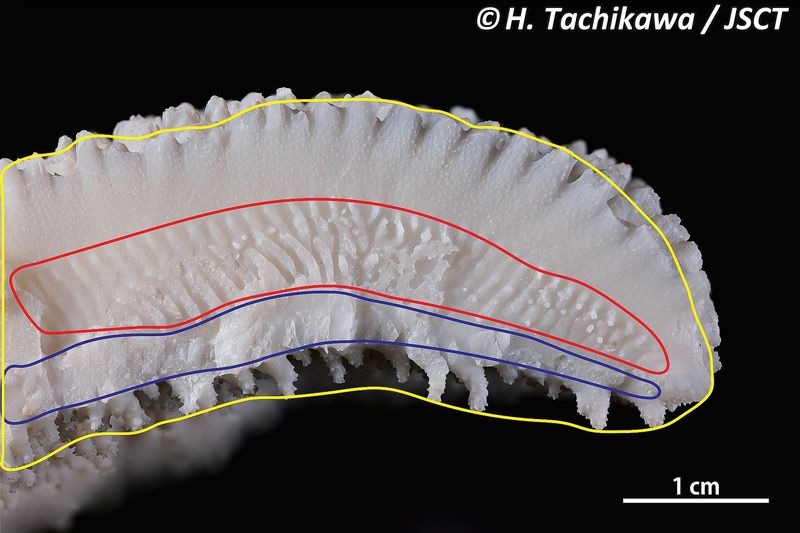 1
1
図1.Ctenactis echinata トゲクサビライシのサンゴ体破断面。 黄色の線で囲んだ部分が一枚の隔壁-肋ユニットで、連続する複合シナプティキュラ (赤色の線で囲んだ部分)
およびサンゴ体壁 (青色の線で囲んだ部分) により隣り合うユニットと結合する。
多くの単口性の種では、大小2種類の隔壁、すなわち大型隔壁と小型隔壁が上面の口を中心に交互に並ぶ。大型隔壁は小型隔壁よりやや突出して厚く、通常サンゴ体の周縁部まで分岐せずに連なる。サンゴ体の成長に伴い、小型隔壁の両側に新しい小型隔壁が形成されて三叉状になり、三叉中央の隔壁は周縁部に向けて高さや厚さが増して新たな大型隔壁となる。このため、隔壁数が増えても大型の隔壁と小型の隔壁が交互に配列する状態は維持される。一部の種では、新たに形成された大型隔壁の内縁の部分が肥厚して上方に突出し、触手葉が形成される
(図2)。本WEB図鑑では、この形態を「大型隔壁の内縁部に触手葉を持つ」と表現する。
 2
2
図2.Lobactis scutaria クサビライシの隔壁配列。
種の識別に使われる形態形質としては、サンゴ体の大きさや形および厚さ、隔壁や肋の次数による相対的な大きさの違いの有無、隔壁の配列が密か疎か、隔壁や肋の縁辺の鋸歯や突起の形状や大きさ・密度、隔壁側面の顆粒の配列様式、サンゴ体壁の穴の有無、自由生活性か固着性か、多口性のものでは二次口の密度や分布様態などが重要である。一部の種は生時の触手の形態に特徴を持つ。近年は分子系統解析の結果に基づく分類の改変が進められ、形態のみでは定義のできない属が少なからず生じている。このような例については、該当する個々の分類群のページで記述する。
生活環:クサビライシ科の模式的な生活環を図3に示した。本科の生活環には以下に記すような様々なパターンが含まれる。
1.単口性・自由生活性の種に典型的にみられる基本的な生活環 (A→B→C→D→E→F→A):有性生殖で生み出されたプラヌラが着底して芽体となってキノコ型に成長し、キノコの傘の部分が自切して自由生活体となり、自由生活体が成熟すると有性生殖でプラヌラが生産される。
1a.芽体柄部からの傘の再生による無性生殖 (F→G→E→F):1の生活環で傘が脱落した芽体の柄の部分から傘が再生し (図4)、成長すると傘が脱落して自由生活体が生み出される。
1b.芽体柄部に生じた複数の傘の癒合を伴う無性生殖 (F→H→I→J):1a の変形で、芽体の柄の部分に複数の傘が形成され、これらが部分的に癒合・一体化後に自切し、複数の口を持つ変形した自由生活体
(図5) が形成される (クローンであるため、癒合・一体化は容易に起こる)。長円形のサンゴ体を持つ種では、この過程を経て三つ叉や四つ叉などのサンゴ体が形成されることがある。
1c.損傷を受けた親サンゴ体からの無性生殖 (F→K→E→F):自由生活性のサンゴ体の一部 (特に口を含む部分) が埋没など何らかの理由で損傷を受けると、生き残った部分に新しい芽体が形成されて
(図6)、新たな自由生活性のサンゴ体が形成される。
1d.縦分裂による無性生殖 (F→L→M):自由生活性のサンゴ体が自切(縦分裂) して扇形の破片となり、それぞれの破片が新たな個体となる、いわゆる“ワレクサビライシ型”の無性生殖
(図7)。
2.多口性・自由生活性の種の生活環 (A→B→C→D→E→F→O→A):プラヌラ由来の芽体から生み出された単口の自由生活体に二次口が形成されて多口となり
(または芽体の段階で二次口が形成されて多口となってから自切し)、多口となった自由生活体が成熟すると有性生殖でプラヌラが生産される。
3.固着性の種の生活環 (A→B→C→D→A/A→B→C→D→N→A):プラヌラに由来する芽体が固着したままで成長し、成熟すると有性生殖でプラヌラが生産される。単口性の種では初口一つのままで成熟するが、多口性の種では成熟前に二次的な口が形成されて多口となる。クサビライシ科以外の多くの固着性有藻性イシサンゴ類と類似の生活環である。
3a.固着性・多口性の種の破片化による自由生活サンゴ体の形成 (N→O):固着性・多口性の種のサンゴ体が、物理的な力で基板から脱落したり破片形成が起こったりすることにより、自由生活のサンゴ体が形成される。
 3
3
図3.クサビライシ科イシサンゴ類の生活環の模式図。 A: プラヌラ幼生;B-D:芽体の初期成長;E: 成長した芽体における傘と柄部の間への自切面の形成;F:
自切により生じた自由生活体と、自由生活体分離後の芽体の柄部;G: 柄部への傘の再形成;H: 柄部への複数の傘の再形成;I, J: 複数の傘の癒合による多口自由生活体の形成;K:軟組織が部分的に損傷した自由生活体上への芽体の再形成;L,
M:縦分裂による複数の自由生活体の形成;N: 触手環内出芽による固着性・多口性サンゴ体の形成;O: 触手環内出芽による自由生活性・多口性サンゴ体の形成。実線は有性生殖サイクル内の段階を、破線は様々な無性生殖の様態を示す。(Hoeksema
1989 を参考に改編)
 4
4  5
5  6
6  7
7
図4.自由生活体分離後の柄部から新たに傘部が形成された Danafungia horrida ノコギリクサビライシ のものと思われる芽体 (図2のGに相当)。
図5.二つの傘の癒合により多口となった Cycloeris cyclolites マンジュウイシのサンゴ体(図2のJに相当)。
図6.大部分の軟組織が損傷した Lobactis scutaria クサビライシのサンゴ体の縁辺部に形成された多数の芽体 (図2のKに相当)。
図7.縦分裂により複数のサンゴ体が分離される途中の Cycloseris fragilis マンジュウイシ属の一種のサンゴ体 (図2のLに相当)。
分類の変遷: 現在クサビライシ科に含められている種のうち最も早く命名されたものは、Linnaeus (1758) による Madrepora fungites (=Fungia fungites) および M. pileus (=Halomitra pileus) であり、その後、多くの著者により多数の属・種タクソンが設立された。19世紀半ばになり、Dana (1846) によりクサビライシ科 Fungiidae
が提唱され、Fungia、Herpetolithus (=Herpolitha)、Halomitra、Polyphyllia、Zoopilus などの自由生活性の諸属が整理された。なお、彼の Fungiidae には、 Pavonia (=Pavona)、Agaricia、Psammocora など現在他の科に置かれている固着性・群体性でサムナステロイド型のサンゴ個体配列を持つ属も含まれていた。
20世紀以降、クサビライシ科の分類について後の研究者に大きな影響を与えた研究がいくつか挙げられる。これらの研究についての概要を以下に列挙するとともに、クサビライシ科の属レベルの分類の変遷を表1に示した。
表1.クサビライシ科における属レベルの分類の変遷。
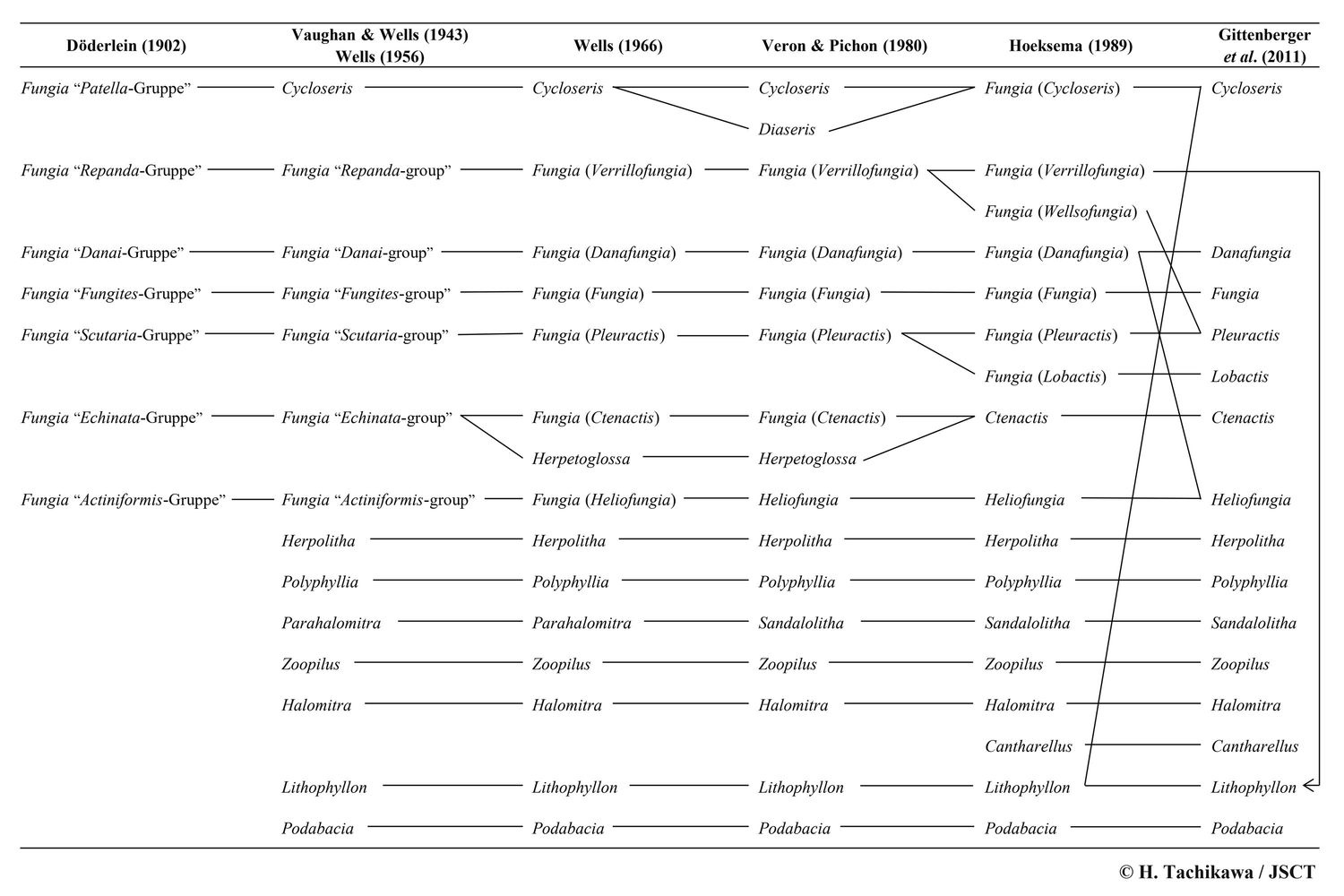
クサビライシ科に関する注目すべき分類学的再検討の最初のものは Döderlein (1902) による Fungia 属の研究であると思われる。彼は、当時単口性・自由生活性のすべての種が含まれていた Fungia 属を、形態に基づき7つの種群に分けた。
イシサンゴ目全体の分類学的再検討を行った Vaughan & Wells (1943) は、Döderlein (1902) による
Fungia 属内の7種群のうち Patella 種群を独立した属 Cycloseris としたが、その他6種群は踏襲した。また、Döderlein (1902) の扱わなかった多口性および固着性の属として7属を認めた。Wells
(1956) はこれと同じ分類体系を示した。なお、Döderlein (1902) から Wells (1956) まで用いられていた「種群」は、動物命名規約の規定するタクソンの学名とは認められない
(命名規約の条4.1の「種階級群よりも高い階級のタクソンの学名は、一語よりなる」を満たさないため)。
Wells (1966) は、Vaughan & Wells (1943) の Fungia 属内の6種群に亜属名を与え、これらのグループが命名規約上で正式なタクソンとして扱われることとなった。また、従来 Echinata 種群に含まれていた多口性の種 Fungia crassa (=Ctenactis crassa) を独立した属とし、新属 Herpetoglossa を設立した。
Veron & Pichon (1980) は、有藻性イシサンゴ類全般にわたるモノグラフ集の中で、クサビライシ科について下記のような改変を行った。
・Cycloseris のシノニムとされていた Diaseris を独立した属とした。
・Fungia (Heliofungia) を属のランクとし、Heliofungia とした。
・Parahalomitra を Sandalolitha のジュニアシノニムとし、後者を有効名とした。
Hoeksema & Best (1984) は固着性・単口性の新属 Cantharellus を提唱し、さらに Hoeksema (1989) はクサビライシ科全般に関する詳細な分類学的再検討を行った。Hoeksema (1989) による主な改変内容は下記のとおりである。
・Cycloseris を、Fungia の亜属とし、Fungia に7亜属を認めた。
・Veron & Pichon (1980) が Cycloseris から分離した Diaseris を再び Cycloseris のシノニムとした。
・Fungia (Verrillofungia) の一部を Fungia (Wellsofungia) とした。
・Fungia (Pleuractis) の一部を Fungia (Lobactis) とした。
・Fungia (Ctenactis) を属のランクとし、Ctenactis とした。
・Herpetoglossa を Ctenactis のシノニムとした。
Gittenberger et al. (2011) は、分子系統解析の成果を盛り込み、下記のような大幅な属レベルの改変を行った。
・Fungia の亜属をすべて属のランクに格上げした。
・Fungia (Wellsofungia) を Pleuractis のシノニムとした。
・Fungia (Verrillofungia) と Lithophyllon は単一のクレードであるとし、後者を有効名とした。
・Fungia (Danafungia) の一部を Heliofungia に編入した。
・Lithophyllon の一部を Cycloseris に編入した。
これらの改変により、Cycloseris と Lithophyllon はそれぞれ単口性・自由生活性の種と多口性・固着性の種の両方を含むこととなり、形態による定義が困難となった。
その後、Benzoni et al. (2012) は分子データに基づきアミメサンゴ科の Psammocora explanulata およびヤスリサンゴ科の Coscinaraea wellsi を Cycloseris に編入した。また、Oku et al. (2017) は Cycloseris hexagonalis に対し、新属 Sinuorota を提唱した。
以上の経緯により、現在本科には16属約58種が含まれ (WoRMS:化石のタクソンを除く)、有藻性イシサンゴ類の中ではサザナミサンゴ科の24属に次ぐ属の多様性が高い科となっている。
タイプ属:Fungia Lamarck, 1801 シタザラクサビライシ属 (原指定によるタイプ)
引用文献:
Benzoni F, Arrigoni R, Stefani F, Reijnen BT, Montano S, Hoeksema B (2012) Phylogenetic position and taxonomy of Cycloseris explanulata and C. wellsi (Scleractinia: Fungiidae): lost mushroom corals find their way home. Contrib Zool 81: 125–146. [ResearchGate]
Dana JD (1846, 1849) United States exploring expedition during the years 1838, 1839, 1840, 1841, 1842 under the command of Charles Wilkes, U.S.N. Vol. VII. Zoophytes. Lea and Blanchard, Philadelphia. [Smithson Lib: text, plates]
Döderlein L (1902) Die Korallengattung Fungia. Abhandlungen herausgegeben von der senckenbergischen naturforschenden Gesellschaft 27: 1–162, I–XXV. [BHL]
Gittenberger A, Reijnen BT, Hoeksema BW (2011) A molecularly based phylogeny reconstruction of mushroom corals (Scleractinia: Fungiidae) with taxonomic consequences and evolutionary implications for life history traits. Contrib Zool 80: 107-132 [ResearchGate]
Hoeksema BW (1989) Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia Fungiidae). Zool Verh 254: 1-295. [ResearchGate]
Hoeksema BW, Best MB (1984) Cantharellus noumeae (gen. nov., spec. nov.), a new scleractinian coral (Fungiidae) from New Caledonia. Zool Meded 58: 323-328. [Naturalis Repository ]
Linnaeus C (1758) Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Vol. 1. Impensis Direct. Laurentii Salvii, Holmiae [BHL]
Loya Y, Sakai K, Heyward A (2009) Reproductive patterns of fungiid corals in Okinawa, Japan. Galaxea J Coral Reef Stud 11: 119-129. [J-Stage]
Oku Y, Naruse T, Fukami H (2017) Morpho-molecular evidence for polymorphism in the mushroom coral Cycloseris hexagonalis (Scleractinia: Fungiidae), with a new phylogenetic position and the establishment of a new genus for the species. Zool Sci 34: 242-251 [ResearchGate]
Vaughan TW, Wells JW (1943) Revision of the suborders, families and genera of the scleractinia. Special papers of the Geological Society of America 44. Waverly Press, Baltimore, MD. [GeoScienceWorld]
Veron JEN, Pichon M (1980) Scleractinia of eastern Australia, part III. Families Agariciidae, Siderastreidae, Fungiidae, Oculinidae, Merulinidae, Mussidae, Pectiniidae, Caryophylliidae, Dendrophylliidae. Australian Institute of Marine Science, Townsville. [BHL]
Wells JW (1956) Part F. Coelenterata: Scleractinia corals. In: Moore RC (ed) Treatise of invertebrate paleontology. Geological Society of America, NY, pp 328-444
Wells JW (1966) Evolutionary development in the scleractinian family Fungiidae. In: Rees WJ (ed) The Cnidaria and their evolution. Symposium of the Zoological Society of London Academic Press, London. 16: 223–246, pl. 1.
執筆者:立川浩之
Citation:
更新履歴:
2024-11-15 公開